please help me with these questions: Describe the DNA-binding domains of four major families of transcription...
please help me with these questions:
- Describe the DNA-binding domains of four major families of transcription factors and their interaction with DNA.
- Describe how CTD phosphorylation affects transcription.
Thanks
Homework Answers
(1) SOLUTION :
Transcription factors are modular proteins composed of distinct and seperable functional domains:
- DNA binding domain (DBD) interacts with specific DNA sequences
- Transcription Activation domain (TAD) interacts with other proteins (general transcription machinery, coactivators etc., to stimulate transcription from the core promoter
Transcription factors that bind DNA do so in a sequence specific manner.
The three common features most DNA binding proteins are:
- They bind to the major groove (12Å wide and 8Å deep ) of B- DNA through α –helices.
- The minor groove of B-DNA (5Å wide, 8Å deep) is generally too narrow to fit entire α –helices.
- They generally do not disrupt the base pairs of the DNA, but distort the conformation of the backbone by bending the double helix
DNA binding domains of eukaryotic transcription factors
In general, transcription factors which bind DNA in a sequence-specific manner contain characteristic DNA binding domains or motifs.Infact, eukaryotic transcription factors are categorized into distinct families based on the type of DNA binding motifs they contains the following
Eukaryotic transcription factor families
- Helix-Turn-Helix proteins
- Zinc finger proteins
- Leucine zipper protein
- Helix-Loop-Helix proteins
1) Helix-turn-Motif
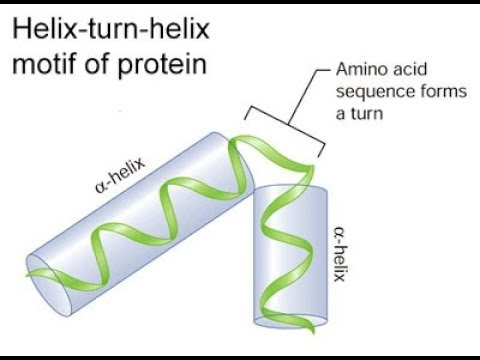
The helix-turn-helix motif was initially identified as the DNA-binding domain of bacteriophage repressors.These repressors contain two alpha helices: one that lies in the major groove of DNA and the other lies at an angle across DNA.Two adjacent α-helices separated by a "turn" of several amino acids enables the protein to bind to DNA . However, the HTH motif cannot fold or function alone, but is always part of a larger DNA-binding domain and amino acid residues outside the HTH motif are important in regulating DNA recognition and binding. A motif similar to helix-turn-helix motif is present in certain eukaryotic transcription factors that play a key role in regulation of embryonic development.The DNA binding domain in these proteins is called the homeodomain and the C-terminal region of homeodomain shows homology with the helix-turn-helix motif of procaryotic repressors. it contains 60 amino acid motif known as homeobox.

Fig1.The λ repressor of bacteriophage lambda employs two helix-turn-helix motifs (left; green) to bind DNA (right; blue and red). The λ repressor protein in this image is a dimer
2) zinc finger motif

Certain eukaryotic transcription factors contain a unique DNA binding motif called a zinc-finger where a zinc ion is coordinated by 2 Cysteine and 2 Histidine residues.This motif was first discovered in a transcription factor known as TFIIIA in the frog, Xenopus laevis.The number of zinc fingers vary in different transcription factors.The non-conserved aminoacids in the C-terminal side of each finger recognize specific target sites.The C-terminal part of each finger forms a-helices that bind DNA while the N-terminal part form b-sheets.A C2H2 zinc finger protein has a series of zinc fingers and the consensus sequence of a single finger is:
Cys-X2-4-Cys-X3-Phe-X3-Leu-X2-His-X3-His
The interspersed cysteine and histidine residues covalently bind zinc atoms, folding the amino acids into loops known as zinc fingers.Each finger consists of approximately 23 amino acids (with a loop of 12 to 14 amino acids between the Cys and His residues) and a linker between loops consisting of 7 or 8 amino acids. The amino acids in the loop bind to specific DNA sequences.
MSNLPPTFGSTRQSPEDQSPPVPKELSFNGTTPSGKLRLFVCQTCTRAFARQEHLKRHER 60
SHTKEKPFSCGICSRKFSRRDLLLRHAQKLHSNCSDAAITRLRRKATRRSSNAAGSISGS 120

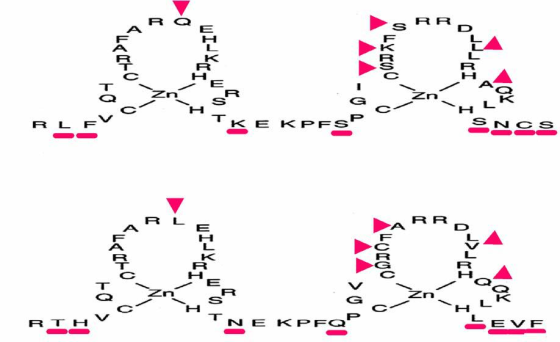
Zinc-finger proteins, represent one of the major families of eukaryotic transcription factors including those belonging to the steroid hormone receptor super family.The steroid hormone receptors contain C2C2 zinc fingers instead of C2H2 fingers. They have the consensus sequence:
Cys-X2-Cys-X13-Cys-X2-Cys

In the case of steroid hormone receptors, the first zinc finger determines specificity of DNA binding and the second finger specifies dimerization specificity.It has been shown that a single amino acid change in the base of the first finger can alter the DNA binding specificity of steroid hormone receptors.
3) Basic leucine zipper (bzip)

In certain transcription factors the dimerization domain forms motif known as leucine zipper.This motif consists of two amphipathic helices, one from each subunit, interacting with each other resulting in a left handed coiled-coil structure.The leucine zipper is a interdigitation of regularly spaced leucine residues in one helix with leucines from the adjacent helix.Leucine zipper motifs can mediate homodimer or heterodimer formation.An important point to note is that the leucine zipper motif itself does not play a directly participate in DNA binding.The DNA-binding domain is adjacent to the leucine zipper and the latter facilitates protein-protein dimerization. Dimerization allows the juxtaposition of the DNA-binding regions of each subunit. A leucine zipper forms an amphipathic helix in which the leucines of the on one protein protrude from the alpha helix and interdigitate with the leucines of the other DNA protein in parallel to form a coiled coil domain.The region adjacent to the leucine repeats is highly basic and forms the DNA-binding site.
4) helix-loop-helix (HLH)

The amphipathic helix-loop-helix (HLH) motif was identified in certain transcription factors involved in developmental regulation. They contain a stretch of 40-50 aminoacids comprising of two amphipathic a-helices separated by a linker region (the loop) of varying length.The HLH proteins form both homodimers and heterodimers by means of interactions between the hydrophobic residues on the corresponding faces of the 2 helices. The ability to form dimers resides with these amphipathic helices.Most HLH proteins contain a basic region adjacent to the HLH motif that is involved in DNA binding and hence they are often called bHLH proteins.Examples of HLH proteins include MyoD, Myf5, myogenin and MRF4 which are involved in myogenesis and are called myogenic regulatory factors (MRFs)
(2) SOLUTION :
1) C-terminal domain (CTD)

Fig : RNA Pol II in action, showing the CTD extension to the C-terminal of POLR2A.
The carboxy-terminal domain (CTD) of RNA polymerase II is that portion of the polymerase that is involved in the initiation of DNA transcription, the capping of the RNA transcript, and attachment to the spliceosome for RNA splicing. The CTD typically consists of up to 52 repeats (in humans) of the sequence Tyr-Ser-Pro-Thr-Ser-Pro-Ser.The carboxy-terminal repeat domain (CTD) is essential for life. Cells containing only RNAPII with none or only up to one-third of its repeats are inviable.The CTD is an extension appended to the C terminus of RPB1, the largest subunit of RNA polymerase II. It serves as a flexible binding scaffold for numerous nuclear factors, determined by the phosphorylation patterns on the CTD repeats. Each repeat contains an evolutionary conserved and repeated heptapeptide, Tyr1-Ser2-Pro3-Thr4-Ser5-Pro6-Ser7, which is subjected to reversible phosphorylations during each transcription cycle.This domain is inherently unstructured yet evolutionarily conserved, and in eukaryotes it comprises from 25 to 52 tandem copies of the consensus repeat heptad.As the CTD is frequently not required for general transcription factor (GTF)-mediated initiation and RNA synthesis, it does not form a part of the catalytic essence of RNAPII, but performs other functions.
2) CTD phosphorylation
RNAPII can exist in two forms: RNAPII0, with a highly phosphorylated CTD, and RNAPIIA, with a nonphosphorylated CTD.Phosphorylation occurs principally on Ser2 and Ser5 of the repeats, although these positions are not equivalent. The phosphorylation state changes as RNAPII progresses through the transcription cycle: The initiating RNAPII is form IIA, and the elongating enzyme is form II0. While RNAPII0 does consist of RNAPs with hyperphosphorylated CTDs, the pattern of phosphorylation on individual CTDs can vary due to differential phosphorylation of Ser2 versus Ser5 residues and/or to differential phosphorylation of repeats along the length of the CTD.The PCTD (phosphoCTD of an RNAPII0) physically links pre-mRNA processing to transcription by tethering processing factors to elongating RNAPII, e.g., 5′-end capping, 3′-end cleavage, and polyadenylation.Ser5 phosphorylation (Ser5PO4) near the 5′ ends of genes depends principally on the kinase activity of TFIIH (Kin28 in yeast; CDK7 in metazoans).The transcription factor TFIIH is a kinase and will hyperphosphorylate the CTD of RNAP, and in doing so, causes the RNAP complex to move away from the initiation site. Subsequent to the action of TFIIH kinase, Ser2 residues are phosphorylated by CTDK-I in yeast (CDK9 kinase in metazoans). Ctk1 (CDK9) acts in complement to phosphorylation of serine 5 and is, thus, seen in middle to late elongation.CDK8 and cyclin C (CCNC) are components of the RNA polymerase II holoenzyme that phosphorylate the carboxy-terminal domain (CTD). CDK8 regulates transcription by targeting the CDK7/cyclin H subunits of the general transcription initiation factor IIH (TFIIH), thereby providing a link between the mediator and the basal transcription machinery.The gene CTDP1 encodes a phosphatase that interacts with the carboxy-terminus of transcription initiation factor TFIIF, a transcription factor that regulates elongation as well as initiation by RNA polymerase II.Also involved in the phosphorylation and regulation of the RPB1 CTD is cyclin T1 (CCNT1).Cyclin T1 tightly associates and forms a complex with CDK9 kinase, both of which are involved in the phosphorylation and regulation.
ATP + [DNA-directed RNA polymerase II] <=> ADP +
[DNA-directed RNA polymerase II] phosphate : catalyzed by CDK9 EC
2.7.11.23.
TFIIF and FCP1 cooperate for RNAPII recycling. FCP1, the CTD
phosphatase, interacts with RNA polymerase II. Transcription is
regulated by the state of phosphorylation of a heptapeptide
repeat.The nonphosphorylated form, RNAPIIA, is recruited to the
initiation complex, whereas the elongating polymerase is found with
RNAPII0. RNAPII cycles during transcription. CTD phosphatase
activity is regulated by two GTFs (TFIIF and TFIIB). The large
subunit of TFIIF (RAP74) stimulates the CTD phosphatase activity,
whereas TFIIB inhibits TFIIF-mediated stimulation.
Dephosphorylation of the CTD alters the migration of the largest
subunit of RNAPII (RPB1).
- 5' capping
The carboxy-terminal domain is also the binding site of the cap-synthesizing and cap-binding complex. In eukaryotes, after transcription of the 5' end of an RNA transcript, the cap-synthesizing complex on the CTD will remove the gamma-phosphate from the 5'-phosphate and attach a GMP, forming a 5',5'-triphosphate linkage. The synthesizing complex falls off and the cap then binds to the cap-binding complex (CBC), which is bound to the CTD.
The 5'cap of eukaryotic RNA transcripts is important for binding of the mRNA transcript to the ribosome during translation, to the CTD of RNAP, and prevents RNA degradation.
- Spliceosome

Fig : Spliceosomal splicing cycle
The carboxy-terminal domain is also the binding site for spliceosome factors that are part of RNA splicing. These allow for the splicing and removal of introns (in the form of a lariat structure) during RNA transcription.
- Mutation in the CTD
Major studies in which knockout of particular amino acids was achieved in the CTD have been carried out. The results indicate that RNA polymerase II CTD truncation mutations affect the ability to induce transcription of a subset of genes in vivo, and the lack of response to induction maps to the upstream activating sequences of these genes.
Add Answer to:
please help me with these questions:
Describe the DNA-binding domains of four major families of
transcription...
Can someone please help me answer these questions. Thank you! Eukaryotic transcription signals a) This drawing shows the placements of the four main sequences of the eukaryotic core promoter for RNA...
 Can someone please help me answer these questions. Thank
you!
Eukaryotic transcription signals
a) This drawing shows the placements of the four main sequences of
the eukaryotic core promoter for RNA polymerase II. Identify each
one and give a brief explanation
b) Which sequences are used in a DPE-driven promoter?
c) Which ones are used in a TATA-driven promoter?
d) Please draw and describe the steps as the transcription factors
work with eukaryotic RNA polymerase II to start transcription of...
Can someone please help me answer these questions. Thank
you!
Eukaryotic transcription signals
a) This drawing shows the placements of the four main sequences of
the eukaryotic core promoter for RNA polymerase II. Identify each
one and give a brief explanation
b) Which sequences are used in a DPE-driven promoter?
c) Which ones are used in a TATA-driven promoter?
d) Please draw and describe the steps as the transcription factors
work with eukaryotic RNA polymerase II to start transcription of...
Please help me on these two questions for me? It's immunology! Please!!? 1. Describe and explain...
Please help me on these two questions for me? It's immunology! Please!!? 1. Describe and explain the main classes of complement activity in the service of the host defense? (In a few sentence or a paragraph) 2. Describe the major steps of class I Endogenous pathway presentation? (In a few sentence or a paragraph) Thanks,
need to help w questions In eukaryotes, the addition of a 5' cap and 3' polyadenylation......
 need to help w questions
In eukaryotes, the addition of a 5' cap and 3' polyadenylation... refer to post-translational modifications occur in the cytoplasm refer to modifications of tRNA are regulated in part by the CTD of RNA polymerase II UD A. seunit Star Transcription activators can function at different steps, which of the following is not one of these steps? releasing RNA polymerase from pause recruiting RNA polymerase to the promoter region/ catalyzing addition of the 5 methyl cap...
need to help w questions
In eukaryotes, the addition of a 5' cap and 3' polyadenylation... refer to post-translational modifications occur in the cytoplasm refer to modifications of tRNA are regulated in part by the CTD of RNA polymerase II UD A. seunit Star Transcription activators can function at different steps, which of the following is not one of these steps? releasing RNA polymerase from pause recruiting RNA polymerase to the promoter region/ catalyzing addition of the 5 methyl cap...
Need help on these questions ? 1.) Steroid hormones trigger transcription of target genes by... ...
Need help on these questions ? 1.) Steroid hormones trigger transcription of target genes by... a. activating the release of Ca+2 ions that bind to and activate transcription factors. b. activating a kinase cascade that results in the activation of a transcription factor for a set of genes. c. binding directly to DNA and stimulating transcription. d. binding to nuclear receptors that act as transcription factors for specific genes. 2.) Which type of cell-surface receptor(s), when...
Please answer these questions correctly! 3. Which of the following compounds are major ligands for receptor...
 Please answer these questions correctly!
3. Which of the following compounds are major ligands for receptor tyrosine kinases (RTKs)? E. Growth factors F. Hormones G. Interferons H. Cytokines Explain the mechanism of activation of RTKs in cell: 4. Which domain of the transcription factor ELK-1 is responsible for binding to DNA? A. A B. B C. C D. D Explain mechanism of activation of ELK-1 by MAPK:
Please answer these questions correctly!
3. Which of the following compounds are major ligands for receptor tyrosine kinases (RTKs)? E. Growth factors F. Hormones G. Interferons H. Cytokines Explain the mechanism of activation of RTKs in cell: 4. Which domain of the transcription factor ELK-1 is responsible for binding to DNA? A. A B. B C. C D. D Explain mechanism of activation of ELK-1 by MAPK:
Topic: Electron Transport/Oxidative Phosphorylation Can someone please help me answer the questions above and explain why...
 Topic: Electron
Transport/Oxidative
Phosphorylation
Can someone please help me answer the questions above and
explain why did you pick that answer?
Thanks
Topic: Electron
Transport/Oxidative
Phosphorylation
Can someone please help me answer the questions above and
explain why did you pick that answer?
Thanks
BIOCHEMISTRY HELP ASAP PLEASE!!! Question 7 3 pts The transcription and production of a protein called...
 BIOCHEMISTRY HELP ASAP PLEASE!!!
Question 7 3 pts The transcription and production of a protein called interferon-beta (FNB) is regulated VF-Blevel by a transcription factor called IRF3 which binds to DNA as a dimer of two 30KD Mutation monomeric proteins. The DNA-binding site of RF3 contains the following amino acids. Nere Arg - Lys - lle - Thr. I create four mutants of IRF3 by changing one of the amino acids of AD the DNA-binding site to serine, and then...
BIOCHEMISTRY HELP ASAP PLEASE!!!
Question 7 3 pts The transcription and production of a protein called interferon-beta (FNB) is regulated VF-Blevel by a transcription factor called IRF3 which binds to DNA as a dimer of two 30KD Mutation monomeric proteins. The DNA-binding site of RF3 contains the following amino acids. Nere Arg - Lys - lle - Thr. I create four mutants of IRF3 by changing one of the amino acids of AD the DNA-binding site to serine, and then...
This question has been bothering me, can you please help? Describe in detail all of the...
This question has been bothering me, can you please help? Describe in detail all of the four factors that contribute to the folding of a polypeptide.
can you please help me with these two questions. thank you Explain the second law of...
 can you please help me with these two questions. thank
you
Explain the second law of thermodynamics and how it related to trophic levels and energy pyramids in systems. Use an example. Explain how biotic factors regulate population abundance. Describe what kinds of biotic factors regulate populations. Give an example.
can you please help me with these two questions. thank
you
Explain the second law of thermodynamics and how it related to trophic levels and energy pyramids in systems. Use an example. Explain how biotic factors regulate population abundance. Describe what kinds of biotic factors regulate populations. Give an example.
Could somebody please help me with this questions, please and thanks you Consider a statistical test....
Could somebody please help me with this questions, please and thanks you Consider a statistical test. Is the purpose to describe a sample, describe a population, or use a sample to conclude something about a population? When conducting a hypothesis test, two hypotheses must be stated. What are they?
 Can someone please help me answer these questions. Thank
you!
Eukaryotic transcription signals
a) This drawing shows the placements of the four main sequences of
the eukaryotic core promoter for RNA polymerase II. Identify each
one and give a brief explanation
b) Which sequences are used in a DPE-driven promoter?
c) Which ones are used in a TATA-driven promoter?
d) Please draw and describe the steps as the transcription factors
work with eukaryotic RNA polymerase II to start transcription of...
Can someone please help me answer these questions. Thank
you!
Eukaryotic transcription signals
a) This drawing shows the placements of the four main sequences of
the eukaryotic core promoter for RNA polymerase II. Identify each
one and give a brief explanation
b) Which sequences are used in a DPE-driven promoter?
c) Which ones are used in a TATA-driven promoter?
d) Please draw and describe the steps as the transcription factors
work with eukaryotic RNA polymerase II to start transcription of...
 need to help w questions
In eukaryotes, the addition of a 5' cap and 3' polyadenylation... refer to post-translational modifications occur in the cytoplasm refer to modifications of tRNA are regulated in part by the CTD of RNA polymerase II UD A. seunit Star Transcription activators can function at different steps, which of the following is not one of these steps? releasing RNA polymerase from pause recruiting RNA polymerase to the promoter region/ catalyzing addition of the 5 methyl cap...
need to help w questions
In eukaryotes, the addition of a 5' cap and 3' polyadenylation... refer to post-translational modifications occur in the cytoplasm refer to modifications of tRNA are regulated in part by the CTD of RNA polymerase II UD A. seunit Star Transcription activators can function at different steps, which of the following is not one of these steps? releasing RNA polymerase from pause recruiting RNA polymerase to the promoter region/ catalyzing addition of the 5 methyl cap...
 Please answer these questions correctly!
3. Which of the following compounds are major ligands for receptor tyrosine kinases (RTKs)? E. Growth factors F. Hormones G. Interferons H. Cytokines Explain the mechanism of activation of RTKs in cell: 4. Which domain of the transcription factor ELK-1 is responsible for binding to DNA? A. A B. B C. C D. D Explain mechanism of activation of ELK-1 by MAPK:
Please answer these questions correctly!
3. Which of the following compounds are major ligands for receptor tyrosine kinases (RTKs)? E. Growth factors F. Hormones G. Interferons H. Cytokines Explain the mechanism of activation of RTKs in cell: 4. Which domain of the transcription factor ELK-1 is responsible for binding to DNA? A. A B. B C. C D. D Explain mechanism of activation of ELK-1 by MAPK:
 Topic: Electron
Transport/Oxidative
Phosphorylation
Can someone please help me answer the questions above and
explain why did you pick that answer?
Thanks
Topic: Electron
Transport/Oxidative
Phosphorylation
Can someone please help me answer the questions above and
explain why did you pick that answer?
Thanks
 BIOCHEMISTRY HELP ASAP PLEASE!!!
Question 7 3 pts The transcription and production of a protein called interferon-beta (FNB) is regulated VF-Blevel by a transcription factor called IRF3 which binds to DNA as a dimer of two 30KD Mutation monomeric proteins. The DNA-binding site of RF3 contains the following amino acids. Nere Arg - Lys - lle - Thr. I create four mutants of IRF3 by changing one of the amino acids of AD the DNA-binding site to serine, and then...
BIOCHEMISTRY HELP ASAP PLEASE!!!
Question 7 3 pts The transcription and production of a protein called interferon-beta (FNB) is regulated VF-Blevel by a transcription factor called IRF3 which binds to DNA as a dimer of two 30KD Mutation monomeric proteins. The DNA-binding site of RF3 contains the following amino acids. Nere Arg - Lys - lle - Thr. I create four mutants of IRF3 by changing one of the amino acids of AD the DNA-binding site to serine, and then...
 can you please help me with these two questions. thank
you
Explain the second law of thermodynamics and how it related to trophic levels and energy pyramids in systems. Use an example. Explain how biotic factors regulate population abundance. Describe what kinds of biotic factors regulate populations. Give an example.
can you please help me with these two questions. thank
you
Explain the second law of thermodynamics and how it related to trophic levels and energy pyramids in systems. Use an example. Explain how biotic factors regulate population abundance. Describe what kinds of biotic factors regulate populations. Give an example.
Most questions answered within 3 hours.
-
Where is the error in this code sequence?
String s1 = "Hello";
String s2 = "ello";...
asked 10 months ago -
Financial data for Joel de Paris, Inc., for last year
follow:
Joel de Paris, Inc.
Balance...
asked 10 months ago -
Consider this reaction:
Al2(SO4)3 (aq)+ BaCl3
(aq) Al2Cl6 (aq)- +
3BaSO4(s) . What is the...
asked 10 months ago -
Suppose that Savneet is considering increasing her
recent random sample from 20 car rentals to 40...
asked 10 months ago -
Trucks arrive at an unloading terminal at an average rate of 120
per hour.
Trucks arrive...
asked 10 months ago -
Why are methanol and ethanol completely soluble in water while
octanol is not very little soluble....
asked 10 months ago -
A facilities manager at a university reads in a research report
that the mean amount of...
asked 10 months ago -
When the CuSO4 is rehydrated by adding water to the anhydrous
compound, is this an endothermic...
asked 10 months ago -
A ray of sunlight is passing from diamond into crown glass; the
angle of incidence is...
asked 10 months ago -
A block of mass 0.249 kg is placed on top of a light, vertical
spring of...
asked 10 months ago -
how do the kidneys compensate in the presences of acidosis
a) trigger hyperventilate
b) reserve acid...
asked 10 months ago -
Question 501 pts
The rental rate of capital to the firm increases. Which of the
following...
asked 10 months ago